多指畸形(多余的脚趾)是一种无害的身体结构,常见于缅因猫(图 1)。缅因猫的多指症是由调节基因 ZRS 内名为“海明威突变”(Hw) 的单点突变引起的。该基因负责 Shh 基因的表达并确定趾头数量的形成(Lange et al. 2014)。Hw 突变会导致不同的表型,不同表型之间差别很大(Lange 等人,2014 年;PolyTrak 数据收集)。患有 Hw 突变的猫可能有多达 8 个额外的脚趾,它们可能只是更大一点的露爪,或者变成拇指(所以仍然是 5 根手指),或者只有一个多趾(6 根手指)。因此,多指畸形的表现可能非常小(就像只有一个小结),或者有几个额外的手指。它们通常被称为“手套”爪子,因为额外的“拇指”脚趾使爪子看起来像手套。导致多指畸形的突变作为具有高外显率的常染色体显性性状遗传(Danforth 1947;Lettice et al. 2008;Hamelin 2011),因此可以在育种中轻松选择。迄今为止,在多趾猫中总共发现了三种突变:缅因猫和精灵鲍勃猫的 Hw 突变,以及英国家养短毛猫的UK1和UK2突变(Lettice et al. 2008,Lyons 2009)。肯定存在更多的突变,因为加拿大多趾缅因猫品种既没有Hw突变,也没有UK1或UK2突变(Hamelin 2011)。法国里昂兽医学校的玛丽·阿比特博尔博士和她的同事目前正在努力找出导致加拿大缅因猫多指的突变。
这篇评论是我在2018年秋季PawAcademy G2课程中选定的主题上作为我的“特别项目”撰写的。文本于2020年1 月更新。

简介
缅因猫于 1976 年被猫爱好者协会 (CFA) 接受为一个品种并授予冠军地位。15 年多后,即 1982 年,该品种也获得了国际猫科动物联合会 (FIFe) 的认可。
缅因猫原产于北美新英格兰地区的缅因州。这些猫通过自然选择进化而来,发展出优秀工作猫的所有特征。缅因猫是体型庞大、聪明、熟练的猎手,它们非常适合在严酷的沿海冬季气候中生存。
多指过去和现在仍然是缅因猫品种的一个常见特征。“polydactyly”一词源自希腊语“polydaktylos”(poly = much、many、 daktylos =手指)并描述了与给定物种的野生型相比,人类和动物具有额外的手指和/或脚趾的身体异常。
多指缅因猫最初占基因库的很大一部分(根据 FIFe 品种委员会 MCO 和 Henning Mueller-Rech,2011/2012,40%),因此多指猫是当今大多数缅因猫血统的一部分。但取消表演资格导致许多多指缅因猫被排除在繁殖之外。多指畸形是一种具有高外显率的常染色体显性性状遗传(Danforth 1947;Lettice et al. 2008;Hamelin 2011),因此在育种中很容易被选择。
多指症并不只在缅因猫身上发现。它在北美东海岸(美国和加拿大)以及英国的猫中很常见。Lloyd (1985) 对加拿大东部和美国东北部 35 个种群中的 5000 多只猫进行了检查,发现这些不同种群中的 0-8% 的猫是多指猫。著名作家欧内斯特·海明威在佛罗里达州基韦斯特和古巴的两个家中养了近 200 只猫,其中许多是有六个脚趾的多指猫。如今,海明威故居和博物馆中仍然生活着 40-50 只这些猫的后代 在基韦斯特。在挪威,多指猫通常被称为“skipskatt”(“船猫”)。这些猫被水手们认为是“幸运的吉祥物”,他们还认为这些猫是优秀的捕鼠能手(Harthwell 2001-2010)。
多指畸形对猫的健康没有任何影响(Lange et al. 2014,Hamelin et al. 2017),应被视为该品种的无害表型变异。一般来说,多指并不是一种缺陷,不会对猫造成任何不良影响。但在极少数情况下指甲生长可能会受到影响。当然,过度生长的爪子不仅限于多趾猫,而且额外的脚趾可能比正常脚趾更短,方向也略有不同。尽管如此,不论是不是多趾,所有猫的爪子和指甲,都应该得到妥善照顾,在需要修剪时修剪指甲,就像我们为我们的狗做的那样。多指缅因猫的大爪子经常被比作雪鞋,一些人认为它们有助于在雪上行走。人们还认为,大爪子可以帮助猫更有效地攀爬和狩猎。
如今,多趾缅因猫可以在 CFA、新西兰爱猫公司 (NZCF)、国际猫协会 (TICA) 和猫爱好者联合会 (CFF) 内繁殖和注册。NZCF 是第一个在展览上接受多趾缅因猫的国家,使其有资格与常规脚趾的缅因猫一起获得冠军地位(2009 年 1 月)(PolyTrack Newsletter 2009)。TICA 效仿这一做法,自 2015 年 5 月起接受多趾缅因猫作为冠军(TICA 网页)。FIFe 却反其道而行之,最近(2014 年 1 月)禁止饲养和注册多趾缅因猫。
猫多指症的遗传学
在基因杂志将多趾作为一个研究领域开始扩展研究之前,对多指症的研究已经持续了 100 年以上,有关多指症的出版物可以追溯到 1861 年(Lange 和 Müller 2017)。
丹福斯 (Danforth) 于 1947 年发表了第一篇关于猫多指遗传的论文。他研究了多指猫几代的遗传,以评估额外手指形成的频率。
丹福斯将多指猫与多指猫、多指猫与非多指猫、非多指猫与非多指猫进行了交配。他发现,在每一次交配中,如果父母中至少有一个是多指的,多指猫也会出现在后代中,而在正常足猫之间的交配中,从来不会出现多指后代。丹福斯还发现,在一些交配中,所有后代都是多指的,并认为这可以用父母之一或双方都是多指纯合子来解释。
丹福斯 (1947) 从他的研究中得出结论,多指症是由可变表达的显性基因引起的,他没有发现任何迹象表明该“基因”在纯合时有害或致命。
此外,丹福斯(1947) 在他对 254 只来自两只家养长毛母猫的近亲繁殖的多指小猫的研究中,没有发现多指与其他异常(如裂足或桡骨发育不良)相关的证据。
多指症是由调节基因 ZRS 中的单点突变引起的
今天我们知道,多指症不是由基因突变(“Pd 基因”)引起的,而是由称为 ZRS 的调节元件的单点突变引起的,ZRS 负责表达决定猫爪子数字形成的基因(Lange et al 2014)。这表明猫轴前多指畸形的性质与其他哺乳动物(小鼠和人类)相似,产生肢体特异性表型,但没有其他身体或生理缺陷。
伯斯兰格等人。(2014)和哈梅林等人。(2017) 记录了猫中多指畸形表达的巨大变异性,但得出的结论是这些单点突变对动物福利没有明显的不良后果。
基因可以编码表型。但也有一些调控基因控制其他基因的表达,并以这种方式影响表型。调控基因可以与 DNA 启动子结合并阻止 DNA 的复制,从而使编码的蛋白质不表达,从而阻止其功能。
如上所述,家猫(以及人类和小鼠)的多指症是由称为 ZRS 的调节基因的单点突变引起的。ZRS 是 SHH(一种称为“Sonic Hedgehog”的信号分子)的长距离、肢体特异性顺式调节因子,控制发育中的肢芽中Shh的表达,从而调节数字身份和数量(图 2)。
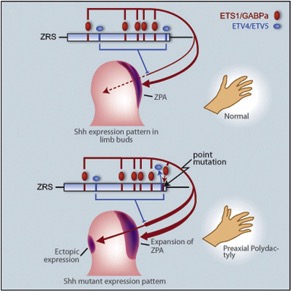
图 2. 调节基因 ZRS 中的单碱基对(点)突变已被证明会导致多指症,因为它们调节肢芽(极化活动区 (ZPA))中的 Shh 表达,并通过异位诱导形成额外的手指Shh 的表达(Lettice et al. 2003;图转载自 Lettice et al. 2012)。
莱蒂斯等人。(2008) 在北美和英国猫的 ZRS 中发现了与轴前多指相关的三种新突变(单核苷酸取代)(图 3)。这些是迄今为止与猫多指相关的唯一已知突变。
多趾海明威的猫(图 3B)要么是杂合子(A/G)(n=3),要么是纯合子(G/G)(n=1)。纯合猫没有表现出更严重的表型,并且仅在前肢上呈多指状。Lettice 等人纳入这项研究的其他美国猫。(2008) (图 3C) 与海明威的猫具有相同的突变,表明北美的多趾猫存在共同的等位基因,而英国猫没有 Hw 突变。相反,英国猫 (n=8) 被发现在位置 257 和 481(UK1 和 UK2)有突变(图 3C 和 D)。
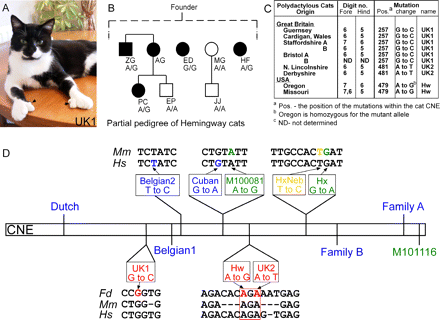
图 3. 多趾猫及其突变的图形和遗传分析,转载自 Lettice 等人。(2008)。 A 显示了一只携带 UK1 突变(第 257 位)的多趾猫,前爪上有多余的手指。B显示了研究中采样的海明威猫的部分血统,所有这些猫都可能来自同一个祖先。受影响的猫用实心正方形(雄性)和圆形(雌性)表示。显示每只猫的姓名缩写,其基因型显示在姓名缩写下方。野生型等位基因在猫序列的第 481 位有一个 A,而突变型等位基因有一个 G. C 显示了来自英国和美国的多趾猫的列表,其中显示了每种猫的起源、在英国猫的位置 257 (UK1) 和 481 (UK2) 以及在位置 479 (Hw) 确定的肢体表型和突变。美国猫。 D 显示了800 bp 保守非编码元件(CNE,它是调节基因 ZRS 中的保守部分/片段)中 12 个突变的总结。猫突变的相对位置以红色显示在 CNE 下方,以及小鼠和人类中的相应序列(Fd, 家猫;Mm, 小家鼠;和 Hs, 智人)。分析中使用的其他突变显示在 CNE 上方(Hs 中识别的突变以蓝色表示,而 Mm 突变以绿色表示)。
猫多指症的基因
检测疾病和表型的 DNA 检测对于世界各地的猫繁育人来说是一项快速增长的有效工具,并且有许多商业实验室和测试可用。基于 Lettice 等人确定的三个已知遗传标记/突变对多指症进行最佳选择 (MyCatDNA) 测试。(2008)。
LIMBR1:c.A479G
单点突变 c.A479G 被 Lettice 等人称为突变 Hw。(2008)(图 3)也被称为“海明威突变”。患有这种突变的猫有 2 到 8 个额外的手指/脚趾。这种突变已在缅因猫(Lettice et al. 2008)和 i PixieBob 猫(Lyons 2009)中发现。
LIMBR1:c.A481T 和 LIMBR1:c.G257C (称为突变 UK2 和 UK1)
Lettice 等人在家养短毛猫中发现了这两种影响基因 Shh的单点突变。(2008)(图3)。
Hw 突变并不像 Lange 等人最初建议的那样是前肢特异性的。
Lange et al.(2008) 宣称猫 Hw 突变是前肢特异性的,并指出海明威的猫主要影响前肢(大约 80% 受影响的猫仅在前爪上表现出多指,参考与 Kirsten Jensen 的个人交流, DVM,佛罗里达州基韦斯特,在海明威故居对海明威猫进行了采样),而携带 UK2 突变的猫的所有爪子上都有额外的手指。同样,莱蒂斯等人。(2008) 描述了 Hw 突变体的独特前肢效应,占 80% 的病例。
我们的缅因猫贝罗娜 (Bellona) 接受了所有三种已知突变变体的检测,并在 ZRS 479 位点的“海明威突变”(杂合 Hw/N)检测呈阳性。她的 DNA 筛查结果是公开的,可在线获取。贝罗娜出生时,每只前爪上都有两个额外的脚趾,每只后爪上都有一个额外的脚趾(7-7-5-5),总共 24 个脚趾。因此,贝罗娜的 Hw 影响了所有四个爪子,并且在她的父亲埃尔德斯和祖母蓝铃花中发现了相同的脚趾模式 - 而她的兄弟伯莎是野生型 (N/N),前爪上有 6 个和 7 个脚趾,每只都有 5 个脚趾。他的后爪(图4)。
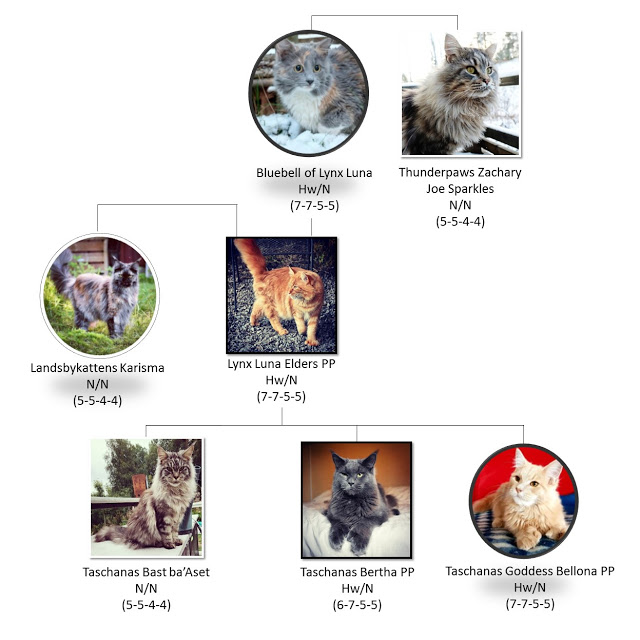
图 4. 我们的多指缅因猫 Taschanas Goddess Bellona PP 的部分族谱。她是“海明威突变”(Hw/N)杂合子,出生时每只前爪上都有两个额外的脚趾,每只后爪上都有一个额外的脚趾(7-7-5-5),24个脚趾总共。贝罗娜的兄弟 (Bertha)、父亲 (Elders) 和祖母 (Bluebell) 都是多指动物。她的另一个兄弟 (Bast)、母亲 (Karisma) 和祖父 (Zack) 都是野生型,有 18 个脚趾。多趾猫用黑色方块(雄性)和圆圈(雌性)表示,野生型猫(N/N)用白色方块(雄性)和圆圈(雌性)表示。摄影:Marita R. Sørensen / Taschanas、Johanna Eli Forssblad / Lynx Luna、Sofie Gonzales、Monica Alterskjær Sundset / Alterskjær。
Lange 等人最近的一项研究。(2014) 表明,在他在这项研究中测试的 378 只多指 Hw 突变猫中,只有 47% 的多指表型仅限于前肢。事实上,缅因猫的 Hw 单点突变被发现会导致许多不同的表型和多达八个额外的脚趾(图 5-6)(Lange 等人,2014 年)。猫也可能有“狼爪”(一种软组织附属物,其爪子缺乏内部骨骼元素)。
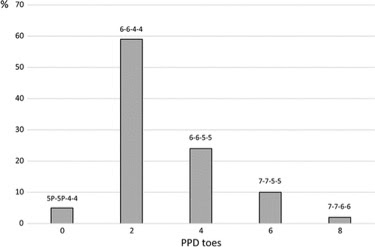
图 5. 猫对称多趾的频率(n=375)随 Hw 稳定,脚趾越多频率越低。所有四个肢体的脚趾数量均显示在柱上方。Hw 突变体中最常见的模式是 6-6-4-4。野生型猫有5-5-4-4个脚趾,总共18个。表中不包括罕见的不对称多指畸形。PPD = 轴前多指畸形,5P = 三指拇指。图转载自 Lange 等人。(2014)。
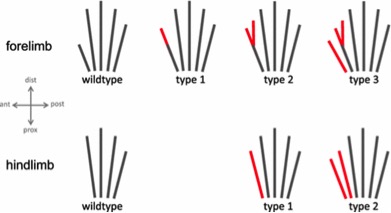
图 6. 该图转载自 Lange 等人。(2014) 并显示了缅因猫的野生型(无多指)和 Hw 突变体肢体模式,其患有轴前多指,指的是爪子内侧边缘(拇指侧)的额外手指(轴后多指指的是外侧(小指侧)有额外的数量,这种情况并不常见)。从上方看到的右前肢(前爪)显示在顶部:1型有一个加长的数字 1;类型 2 具有与类型 1 相同的细长指一,但此外还具有后分叉;类型 3 与类型 2 相同,加上一个完整的额外前指。拇指上有多余手指的爪子(2型和3型)通常被饲养员称为“手套爪”。从上面看到的右后肢(后爪)如图底部所示:
与猫多指相关的更多突变仍有待发现
“小脚”(多出脚趾,但没有“拇指”)(图 1)背后的突变仍有待发现。此外,Hamelin (2011) 在他的博士项目中检测到了他研究中所有多指缅因猫的 Hw 突变,但在加拿大血线的猫中却没有检测到 Hw 突变。重要的是,Hamelin 和同事还排除了这些加拿大猫的 ZRS 或邻近 DNA 区域中存在任何其他突变。这就提出了一个问题:调节基因 ZRS 之外也一定存在突变,这可能导致猫的多指畸形。法国里昂兽医学校的玛丽·阿比特博尔博士和她的同事目前正在努力揭示加拿大血线中的第二个未知突变(个人交流)。
Danforth CH (1947) 猫多指畸形的遗传。J Hered 38: 107-112
Hamelin A (2011) La Polydactylousie du Maine Coon。阿尔福国立兽医学院。论文
Hamelin A、Begon D、Conchou F、Fusellier M、Abitbol M (2017) 缅因猫多指畸形的临床特征。猫科动物医学和外科杂志 19:382-393。
Harthwell S (2001-2010) 多趾猫(第 1 部分)。
Lange A、Nemeschkal HL、Müller GB (2014) 携带单点突变的多指猫的偏向多型现象:数字新颖性的海明威模型。进化生物学四十一:262-275。
兰格 A,穆勒 GB (2017) 多指症的发展、遗传和进化。生物学季刊 92(1)。
Lettice LA, Heaney SJH, Prudie LA,Li L, de Beer P, Oostra BA, Goode D, Elgar G, Hill RE, Gde Graaff E (2003) 长程 Shh 增强子调节发育中的 肢体和鳍的表达,并且是与轴前多指症有关。人类分子遗传学 12:1725-1735
Lettice LA、Hill AE、Devenney PS、Hill RE (2008) 远距离音刺猬顺式调节器中的点突变产生可变的调节输出,导致轴前多指畸形。人类分子遗传学 17:978-985
Lettice LA、Williamson I、WiltshireJH、Peluso S、Devenney PS、Hill AE、Essafi A、Hagman J、Mort R、Grimes G、DeAngelis CL、Hill RE (2012) ETS 因子家族的相反功能定义了唇芽和多指畸形中的 Shh 空间表达。发育细胞 22: 459-467
Lloyd AT (1985) 新英格兰和加拿大滨海地区家猫种群中突变等位基因的地理分布。Journal of Biogeography 12: 315-322
Lyons LA ( 2009) 猫遗传学的最新进展。CAB 评论:农业、兽医科学、营养和自然资源展望 4 (018):1-14
最优选择 (MyCatDNA) – 有关猫科动物遗传育种分析的在线信息。
PolyTrak 时事通讯 – Paws for Effect (2009) NZCF 是世界上第一个接受缅因浣熊毛的大型猫科动物协会!3(1): 3
FIFe 品种委员会 MCO 和 Henning Mueller-Rech,2011 年 2 月/2012 年 2 月,历史上 FIFe 中的缅因浣熊标准以及目前关于 缅因浣熊多指的
TICA
作者: Monica A Sundset 翻译:小娘鱼
")
")
")
")
")
")
")
")
")
")